- Présentation
- Recherche
- DGG : Département Génomique et Génétique du Développement
- REGARN : Les ARN non-codants, des acteurs de la plasticité développementale de la racine
- ChromD : Dynamique des chromosomes
- SILEG : Voies de signalisation contrôlant le développement du système racinaire des légumineuses
- FLOCAD : Développement floral et déterminisme du sexe
- Qlab : Equipe Génomique et épigenomique quantitative des plantes
- DPHYS : Département Physiologie et Signalisation
- PMIN : Département Interactions Plantes Micro-organismes et Réseaux
- DGG : Département Génomique et Génétique du Développement
- Enseignement
- Plateformes
- Bases de données
A la croisée des voies systémiques régulant la nodulation symbiotique
La voie “Compact Root Architecture 2” (CRA2) favorise la compétence des racines à noduler via l’effecteur systémique miR2111
En condition de carence azotée, les légumineuses sont capables de former de nouveaux organes au niveau de leur système racinaire, les nodosités, grâce à une symbiose avec des bactéries fixatrices d’azote, les rhizobia. Comme cette interaction est bénéfique pour la plante hôte, mais également très couteuse en énergie, la nodulation est finement régulée par des voies de signalisation systémiques (à longue distance) afin de limiter ou favoriser les infections par les rhizobia en fonction de précédentes infections ou de la disponibilité en azote. Chez la légumineuse modèle Medicago truncatula, les peptides de signalisation CLE12 (« Clavata3/Embryo Surrounding Region 12 ») et CLE13 produits dans les racines nodulées sont perçus par le récepteur SUNN (« Super Numeric Nodules ») dans les parties aériennes des plantes pour réguler négativement la nodulation. Inversement, les peptides de signalisation CEP (« C-Terminally Encoded Peptides ») sont produits dans les racines en condition de carence azotée et sont perçus par le récepteur CRA2 (« Compact Root Architecture 2 ») dans les parties aériennes des plantes pour favoriser la nodulation avant même que les rhizobia ne soient présents.
Dans leur dernière étude (Gautrat et al 2020, Current Biology), l’équipe SILEG, dirigée par Florian Frugier, a démontré qu’un des effecteurs en aval de ces voies systémiques est le microARN miR2111, produit dans les parties aériennes, et qui régule négativement l’accumulation des transcrits TML1 (Too Much Love 1) et TML2 dans les racines pour finalement promouvoir la nodulation. La carence azotée et les peptides CEP1 induisent, en l’absence de rhizobia, la production du miR2111 de manière dépendante du récepteur CRA2 dans les parties aériennes de la plante, ce qui favorise la compétence des racines à noduler. En parallèle de la voie de signalisation SUNN qui régule négativement l’accumulation du même effecteur systémique, le miR2111, quand les racines sont nodulées, l’ensemble de ces voies systémiques permet de contrôler dynamiquement la capacité des légumineuses à former des nodosités en fonction de la disponibilité en azote et de la présence de nodosités préexistantes.
Contact: florian.frugier@cnrs.fr
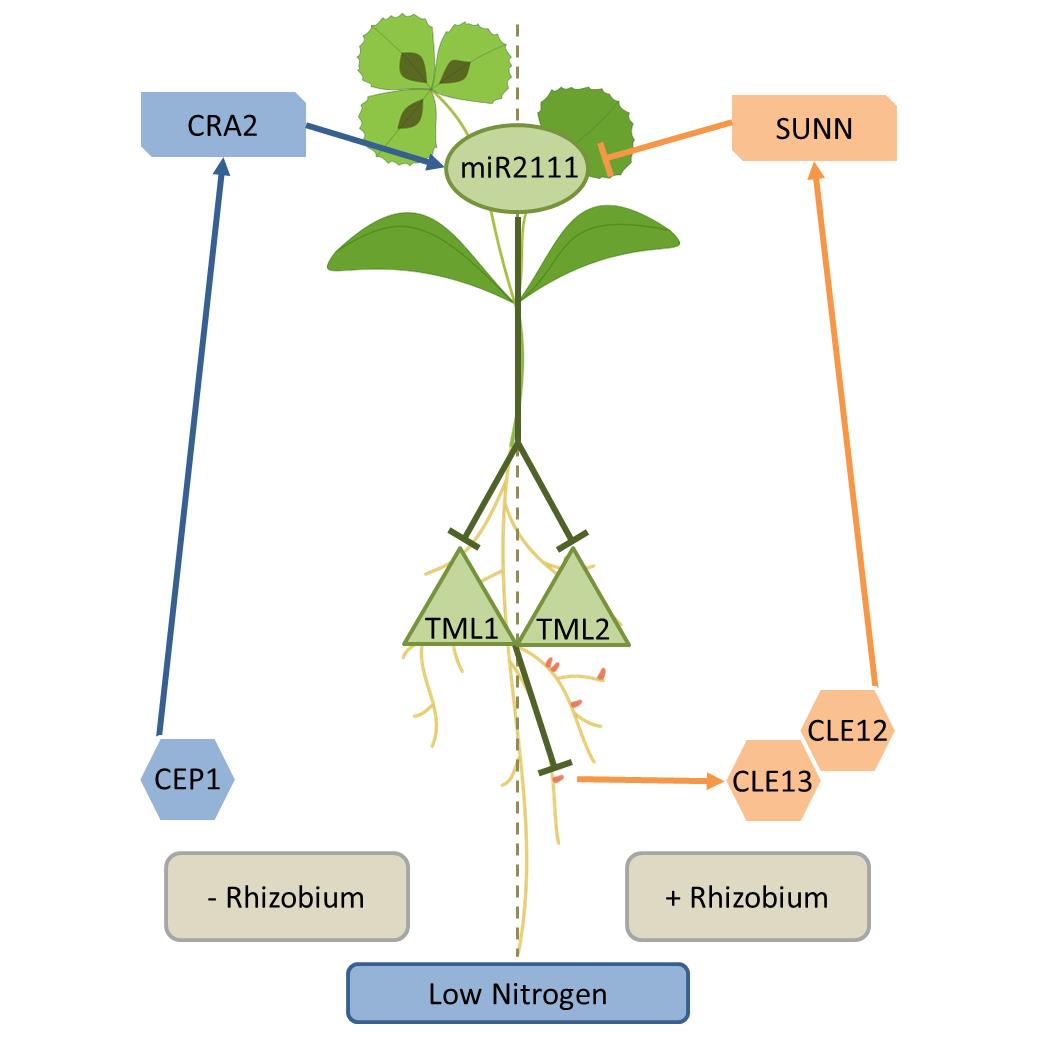
Figure 1. Schéma représentant les voies systémiques régulant la nodulation chez M. truncatula.
En condition de carence azotée, la voie CRA2 régule positivement la nodulation avant même que les rhizobia infectent les racines, via le module régulateur systémique miR2111/MtTML. Après infection par les rhizobia, la voie SUNN régule négativement la nodulation via le même module régulateur systémique. Au final, l’équilibre entre ces voies systémiques permet une régulation fine et dynamique du nombre de nodosités formées par la plante en fonction de la disponibilité en azote et d’infections préexistantes par les rhizobia